Lucas Pedro Cipriani1, Lucas Nascimento Brum1, Rafaela Leopoldina Silva Nunes1, Giovani Luciano Wrasse1, Carine Batista Borges1, Fernando Sintra Fulaneti1, Jessica Deolinda Leivas Stecca1, Thomas Newton Martin1,2
1 Introdução
O crescimento da população mundial aumenta a demanda por alimentos, ao mesmo tempo em que a agricultura enfrenta limitações como a escassez de áreas produtivas, intensificação de eventos climáticos, exigências legais mais rigorosas e a crescente demanda por práticas sustentáveis (Marin et al., 2016). A agricultura intensiva, por sua vez, tem contribuído para a degradação do solo, redução da biodiversidade e contaminação de recursos naturais (Barboza et al., 2012). Além disso, o uso excessivo de agrotóxicos favorece o surgimento de patógenos resistentes.
Nesse contexto, os bioinsumos surgem como alternativa viável para uma agricultura mais sustentável. Bactérias dos gêneros Bradyrhizobium e Azospirillum realizam a fixação biológica de nitrogênio, reduzindo a necessidade de fertilizantes sintéticos e minimizando os impactos ambientais (Prando et al., 2022). Microrganismos utilizados no controle de pragas e doenças ampliam as estratégias de manejo, contribuindo para a redução da resistência de patógenos e promovendo um controle mais eficaz (Chulze, 2023). Além disso, os bioinsumos costumam ter menor custo em comparação aos produtos químicos, sendo economicamente vantajosos ao produtor.
É importante destacar que recursos como fósforo e recursos fosseis utilizados na produção de nitrogênio (ureia) são finitos, e determinadas bactérias e fungos têm se mostrado eficientes na disponibilização desses nutrientes para as plantas. Os microrganismos exercem papel fundamental na dinâmica dos ecossistemas, promovendo alterações nos atributos físicos e químicos do solo. No entanto, seu desenvolvimento depende de condições específicas, como clima, disponibilidade de nutrientes e presença de matéria orgânica (Fuentes-Ramírez et al., 2018; Hawkes et al., 2010). Estratégias de manejo que considerem esses fatores são essenciais para o sucesso da aplicação dos bioinsumos.
Dessa forma, o objetivo deste trabalho é apresentar a relação fisiológica entre microrganismos e as plantas, bem como destacar práticas de manejo indispensáveis para aumentar a eficiência nos cultivos agrícolas.
2. Microrganismos solubilizadores de nutrientes
2.1 Aplicações de bioinsumos solubilizadores na nutrição vegetal
Microrganismos solubilizadores, como bactérias e fungos, desempenham um papel essencial na agricultura ao promoverem transformações bioquímicas que tornam nutrientes minerais mais disponíveis para as plantas. Esses organismos são capazes de solubilizar elementos como fósforo (P) e potássio (K), convertendo-os de formas insolúveis para formas assimiláveis, o que contribui significativamente para a melhoria da fertilidade do solo e o aumento da produtividade agrícola. Além disso, muitos desses microrganismos atuam como agentes de biocontrole, por meio da produção de antibióticos, hormônios vegetais e metabólitos secundários que auxiliam as plantas a tolerar estresses bióticos e abióticos. Com isso, favorecem a redução do uso de fertilizantes químicos e promovem uma agricultura mais sustentável (Olaniyan et al., 2022).
Diversos gêneros bacterianos e fúngicos têm sido relatados como eficazes solubilizadores de fósforo inorgânico, incluindo Bacillus, Micrococcus, Pseudomonas, Burkholderia, Rhizobium, Agrobacterium, Azotobacter, Erwinia, Aspergillus e Penicillium (Rodríguez & Fraga, 1999; Coutinho et al., 2012). Esses microrganismos produzem ácidos orgânicos — como os ácidos láctico, glicólico, cítrico, oxálico, málico, succínico e tartárico — que atuam diretamente sobre os compostos fosfatados, promovendo sua solubilização por meio de mecanismos como a acidificação do meio e a quelação de cátions associados ao ânion fosfato (Andreote & Cardoso, 2016; Mahdi et al., 2011).
No caso do potássio, microrganismos como Acidothiobacillus ferrooxidans, Aspergillus terreus, Bacillus circulans, B. edaphicus, B. mucilaginosus e Paenibacillus spp. apresentam capacidade de solubilizar formas fixas de K presentes em minerais do solo. Essa disponibilidade é promovida por mecanismos como a produção de ácidos orgânicos, formação de biofilmes acidolíticos em superfícies minerais e complexação de íons, favorecendo o crescimento vegetal e o aumento da produtividade das culturas (Maurya et al., 2016).
Diante do crescimento populacional global e da consequente elevação na demanda por alimentos, a pressão sobre os recursos naturais torna-se cada vez maior. Nesse contexto, o uso de microrganismos solubilizadores surge como uma estratégia promissora para enfrentar os desafios da agricultura moderna. Sua inoculação em solos, especialmente em regiões tropicais com baixa fertilidade natural — como os solos brasileiros —, pode reduzir significativamente a dependência de fertilizantes químicos e promover sistemas agrícolas mais resilientes e sustentáveis (Wang et al., 2023).
2.2 Processos microbianos na disponibilização de fósforo no solo
As plantas desenvolveram diferentes mecanismos adaptativos para otimizar a aquisição de fósforo (P), um nutriente essencial, porém frequentemente limitante nos solos agrícolas. Entre essas estratégias, destaca-se a modificação da dinâmica de solubilização e adsorção do fósforo no solo, favorecendo sua difusão até a zona de absorção radicular. Além disso, as plantas podem aumentar a razão raiz/parte aérea, expandir o volume do sistema radicular e elevar a taxa de absorção por unidade de raiz.
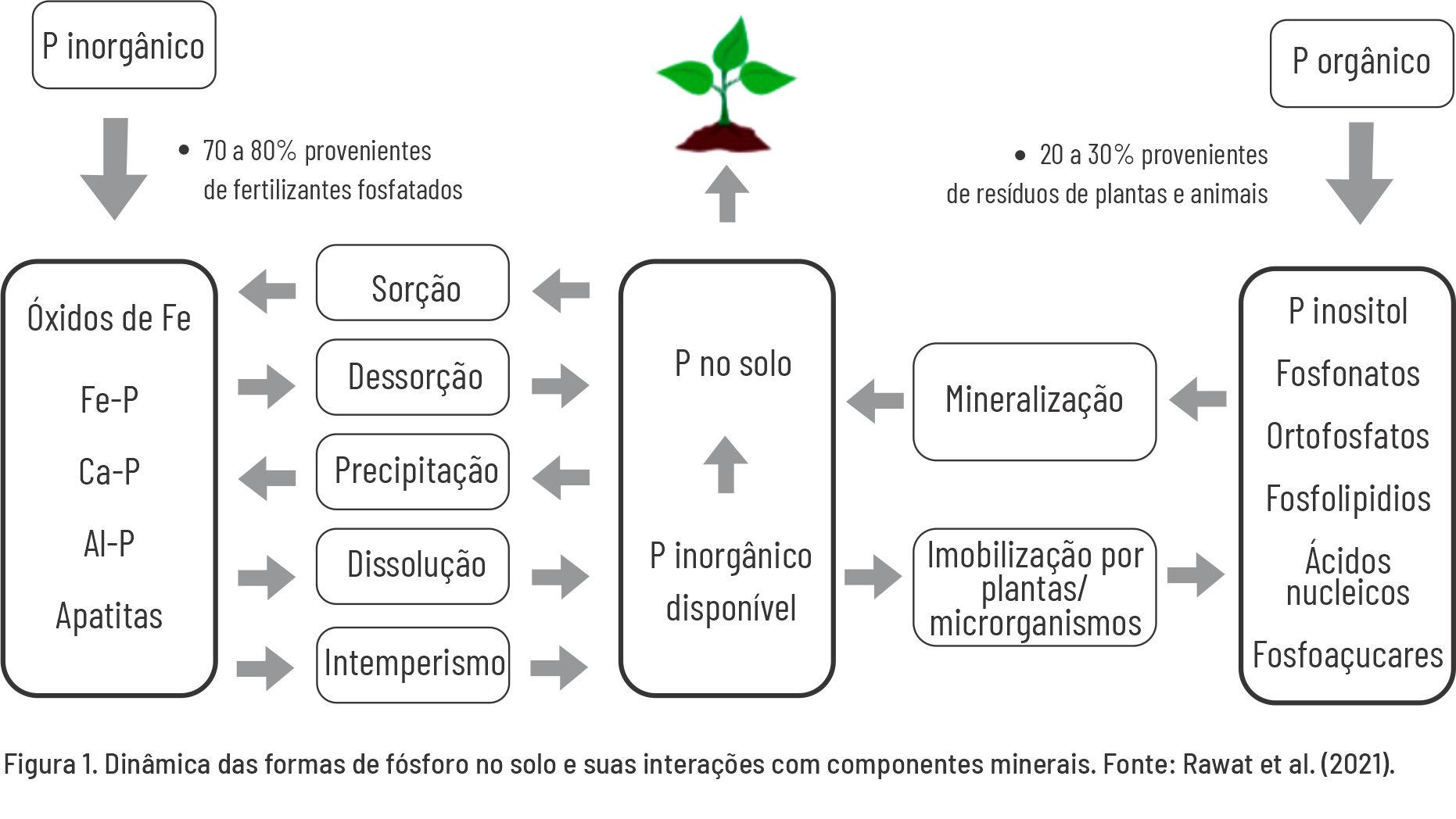
Esses processos são frequentemente acompanhados por uma maior exsudação de fosfatases e outros compostos orgânicos pela rizosfera, bem como pela intensificação da associação com microrganismos micorrízicos (Lajtha & Harrison, 1995). Esses mecanismos indicam que processos geoquímicos e biológicos atuam sinergicamente para transformar os fosfatos naturais do solo em formas mais estáveis — tanto orgânicas quanto inorgânicas. Em solos altamente intemperizados, predominam formas inorgânicas fortemente adsorvidas à fração mineral, com alta energia de ligação, além de formas orgânicas fisicamente e quimicamente estabilizadas (Figura 1).
A produção de ácidos orgânicos tem como base a degradação da glicose no interior das mitocôndrias, resultando na exsudação de compostos como citrato, malato e oxalato. Esses ácidos orgânicos são produtos do metabolismo celular, originando-se da respiração de compostos orgânicos de carbono ou por meio de oxidações diretas — rotas metabólicas primárias envolvidas nesse processo. Durante a inoculação, as bactérias colonizam a superfície das raízes e passam a utilizar glicose proveniente tanto da decomposição da matéria orgânica do solo quanto da exsudação radicular. Nesse contexto, destaca-se a relevância da adição de palhada ao sistema agrícola, a qual contribui para a manutenção de condições favoráveis de temperatura e umidade no solo, além de fornecer fontes adicionais de carbono e açúcares que favorecem a atividade microbiana. Dessa forma, as bactérias solubilizadoras de fósforo presentes na rizosfera produzem ácidos orgânicos capazes de solubilizar o fosfato previamente adsorvido nas frações minerais do solo, especialmente na argila, tornando-o disponível para absorção pelas plantas.
No contexto do mecanismo de acidificação do meio, destaca-se um paralelo com o processo industrial de produção de fertilizantes fosfatados. neste processo, rochas com alto teor de fósforo são extraídas e moídas, sendo posteriormente tratadas com ácidos para promover a liberação do fósforo contido nos minerais. De forma análoga, é fundamental compreender a dinâmica da absorção de nutrientes pelas plantas, que ocorre predominantemente por meio do transporte ativo primário. neste mecanismo, a célula vegetal consome uma molécula de ATP, promovendo a liberação de prótons (H+) para o exterior da célula, contribuindo para a acidificação do meio.
Microrganismos solubilizadores de fósforo (P), como determinadas bactérias e fungos, empregam um mecanismo semelhante. Ao exsudarem prótons no ambiente adjacente às suas células, promovem a acidificação local, favorecendo a dissolução de fosfatos insolúveis presentes na matriz do solo.
Estudos sobre a solubilização de fósforo indicam que a redução do pH no solo está diretamente relacionada ao aumento da disponibilidade de nutrientes. Esse processo envolve a degradação da estrutura mineral ou orgânica que contém o nutriente, seguida pela sua liberação em formas assimiláveis pelas plantas. Para que tal mecanismo ocorra, é essencial que os microrganismos disponham de fontes de carbono, como a glicose proveniente da exsudação radicular ou da decomposição da matéria orgânica do solo.
Durante a respiração celular, a oxidação da glicose nas mitocôndrias leva à geração de ATP e subsequente liberação de prótons (H+), contribuindo para a acidificação do ambiente. portanto, a presença de matéria orgânica no solo é indispensável, pois além de fornecer nutrientes e energia para os microrganismos, mantém condições adequadas de umidade e temperatura, favorecendo sua atividade e eficácia na solubilização de nutrientes.
2.3 Estratégias biológicas para a solubilização de fosfato orgânico
A adição de resíduos de tecidos vegetais e microbianos ao solo, seguida por sua decomposição, constitui a principal origem do fósforo orgânico (Martinazzo et al., 2007). Embora o fosfato orgânico represente, em geral, uma fração menos expressiva do fósforo disponível no sistema solo-planta, sua relevância torna-se mais evidente em ambientes com baixa disponibilidade de fósforo inorgânico (Gatiboni et al., 2008). Estudos demonstram que, em solos não submetidos à adubação mineral, o fósforo orgânico pode contribuir com até 43% do fósforo absorvido pelas plantas, enquanto, em sistemas fertilizados com fontes minerais, essa contribuição reduz-se para aproximadamente 6% (Gatiboni et al., 2005).
A mineralização do fósforo orgânico presente nos resíduos vegetais é promovida, principalmente, pela ação de enzimas como fosfatases e fitases, sintetizadas tanto por plantas quanto por microrganismos. Por meio da hidrólise de compostos orgânicos, essas enzimas liberam fósforo inorgânico na solução do solo, tornando-o disponível para absorção radicular (Nunes, 2011; Gatiboni et al., 2008). No entanto, a eficiência desse processo é condicionada por fatores externos, tais como a quantidade e a qualidade da matéria orgânica presente, além das condições ambientais predominantes (Carvalho et al., 2014). A intensidade da mineralização também está diretamente relacionada à proporção de fósforo orgânico em relação ao total, ao pH do solo e à temperatura (Conte et al., 2002; Nunes, 2011).
Os solos apresentam capacidade de adsorver fósforo; contudo, os sítios de troca são finitos e possuem diferentes graus de afinidade com o íon fosfato (Oliveira et al., 2014). Dessa forma, o potencial de solubilização do fósforo varia conforme as características do solo, sendo que solos argilosos apresentam maior capacidade de retenção do nutriente. Adicionalmente, solos com maior número de sítios de adsorção propiciam condições favoráveis para a ação de bactérias solubilizadoras de fósforo.
O manejo do solo é outro fator determinante na dinâmica do fósforo. Em pastagens naturais cultivadas em solos de baixa fertilidade, a aplicação de fertilizantes fosfatados solúveis em água contribui para o aumento da disponibilidade de frações inorgânicas lábeis, reduzindo a dependência da mineralização do fósforo orgânico (Oliveira et al., 2014). Todavia, após cultivos consecutivos e o consequente esgotamento do fósforo disponível, todas as formas de fósforo no solo podem atuar como fontes tamponantes, liberando fósforo solúvel para as plantas (Santos et al., 2008). Assim, o fósforo orgânico não deve ser negligenciado. Sua contribuição pode ser ainda mais significativa em sistemas de manejo que favoreçam o acúmulo de matéria orgânica, como a fertilização orgânica e práticas conservacionistas (Braos et al., 2015).
O sistema de plantio direto tem se mostrado eficaz na elevação dos teores de matéria orgânica do solo, favorecendo a ciclagem de nutrientes e o acúmulo de formas lábeis de fósforo. Isso ocorre porque os ácidos orgânicos oriundos da decomposição da matéria orgânica atuam na cobertura de óxidos de ferro (Fe) e alumínio (Al), bloqueando seus sítios de adsorção e promovendo a complexação (Zamuner et al., 2008). A ciclagem de nutrientes é essencial para a sustentabilidade dos sistemas produtivos, sendo estimado que entre 77% e 79% do fósforo presente em folhas e raízes mortas possa ser disponibilizado para culturas subsequentes após o cultivo de plantas de cobertura (Borkert et al., 2003). Outro fator de destaque no manejo da disponibilidade de fósforo é a adubação nitrogenada, uma vez que esta promove o aumento da biomassa vegetal, intensificando a deposição de resíduos orgânicos e, consequentemente, a ciclagem de fósforo (Leite et al., 2016).
As fosfatases ácidas não específicas englobam dois principais grupos de enzimas: fosfatases ácidas e fosfatases alcalinas, sendo as primeiras mais relevantes no que se refere à disponibilização do fósforo orgânico no solo. A concentração de fosfatase ácida pode ser utilizada como indicador da atividade microbiana no solo, uma vez que sua expressão tende a aumentar em condições de baixa disponibilidade de fósforo e em sistemas agrícolas submetidos a cultivos sucessivos. Nesses contextos, observa-se um incremento da atividade de organismos exsudadores de fosfatases — como a biomassa microbiana e as plantas — resultando em maior acidificação do ambiente rizosférico e intensificação da competição entre microrganismos e plantas pelo fósforo (Gatiboni et al., 2008).
No manejo agrícola com uso de culturas de cobertura, os microrganismos atuam na degradação de membranas celulares, promovendo a liberação de fósforo orgânico. Em tais sistemas, estima-se que entre 20% e 30% do fósforo presente possa ser absorvido sob a forma de compostos orgânicos. Estudos conduzidos em solos bem manejados, com alta deposição de resíduos vegetais, revelam que o fósforo microbiano constitui um compartimento significativo, contribuindo com a liberação de fosfatos solúveis no sistema. Esses fosfatos, ao serem temporariamente imobilizados, atuam como um mecanismo de retardamento da adsorção do nutriente, sendo mineralizados de forma gradual, o que possibilita um sincronismo mais eficiente com a demanda de absorção das plantas (Conte et al., 2002; Martinazzo et al., 2007).
A atividade das enzimas fosfatases é diretamente influenciada pelo pH do solo. Tanto as fosfatases ácidas quanto as alcalinas apresentam variações de desempenho em função deste parâmetro. O aumento do pH favorece a atividade da fosfatase ácida, o que pode ser atribuído à intensificação da atividade biológica, tornando essa enzima um possível indicador da saúde biológica do solo. No entanto, em solos aditivados com fertilizantes fosfatados, é comum observar uma redução na atividade dessas enzimas, em razão da elevação da concentração de fósforo disponível no meio.
No contexto do ciclo do fósforo no solo, a mineralização do fitato representa um dos processos biológicos centrais e constitui um passo estratégico para a substituição ou complementação da adubação fosfatada convencional por alternativas biológicas. O fitato, ou ácido fítico, corresponde à forma orgânica de fósforo mais abundante presente na matéria orgânica do solo. Sua degradação é mediada por enzimas do grupo das fosfomonoesterases, comumente denominadas fitases, que pertencem à classe das fosfatases. Essas enzimas catalisam a hidrólise das ligações fosfato presentes nas moléculas orgânicas, promovendo a liberação do fósforo na forma solúvel, tornando-o disponível para absorção pelas plantas. As fitases são sintetizadas por diversos organismos, incluindo microrganismos, plantas e tecidos animais (Konietzny & Greiner, 2002).
3. Estratégias microbianas para estimular o crescimento de plantas
A resposta a essa questão está relacionada à produção de fitormônios, que são substâncias reguladoras do crescimento das plantas e podem ser produzidas por plantas, algas e certos microrganismos. Os principais fitormônios e suas funções são: Ácido indolacético (AIA) - Auxinas, responsáveis pelo crescimento e expansão das células; Ácido abcísico (ABA) - Controle hormonal e fechamento estomático; Citocininas - Estímulo à multiplicação celular e à manutenção das folhas verdes; Giberelinas - Estímulo ao crescimento das plantas.
Os fitormônios atuam na planta por meio de sinais químicos que estabelecem uma comunicação entre células, tecidos e órgãos. Cada hormônio vegetal desempenha sua função específica, regulando o metabolismo, o crescimento e a morfogênese da planta (Taiz; Ziegler, 2016). Em muitos casos, dois ou mais fitormônios atuam em conjunto para promover efeitos sinérgicos (Saharan, 2011).
As rizobactérias, por sua vez, habitam as raízes das plantas e exercem efeitos positivos que podem ocorrer tanto de forma direta quanto indireta, promovendo o crescimento vegetal (Kloepper et al., 1980). Diversas espécies de bactérias, como as dos gêneros Pseudomonas, Azospirillum, Azotobacter, Klebsiella, Enterobacter, Alcaligenes, Arthrobacter, Burkholderia, Bacillus e Serratia, foram identificadas com potencial para aumentar o crescimento das plantas (Kloepper et al., 1989; Joseph et al., 2007).
O AIA é uma molécula essencial na sinalização do desenvolvimento da planta, influenciando processos como organogênese, respostas trópicas, expansão celular, divisão e diferenciação celular, além da regulação gênica (Ryu et al., 2008). Vários microrganismos possuem a capacidade de produzir auxinas, como o AIA, por diferentes vias biossintéticas. As interações entre as bactérias produtoras de AIA e as plantas resultam em diversas reações, variando desde a patogênese até a fitoestimulação. As bactérias utilizam o AIA como uma estratégia de colonização, promovendo fitoestimulação e evasão dos mecanismos basais de defesa das plantas (Saharan, 2011).
Os efeitos das auxinas nas plantas são dependentes de sua concentração: em baixas concentrações, podem estimular o crescimento, enquanto em concentrações elevadas, podem ser inibitórias. Assim, diferentes espécies de plantas respondem de maneira distinta às concentrações de auxina (Arshad et al., 1991). As espécies que produzem maiores quantidades de auxinas, como AIA e acetamida indol, promovem o crescimento e o aumento do rendimento da cultura do trigo. Mesmo as cepas que produzem quantidades menores de AIA liberam o hormônio continuamente, o que também contribui para a melhoria do crescimento das plantas (Khalid et al., 2004).
Quanto aos estresses abióticos, como déficit hídrico e temperatura, esses fatores podem prejudicar tanto o crescimento das plantas quanto dos microrganismos. A inoculação com Bacillus aryabhattai demonstrou ser eficaz na mitigação dos efeitos do estresse hídrico e térmico em plantas de soja. Estudos indicaram que a inoculação com essa bactéria promoveu o crescimento das plantas de soja sob diferentes regimes de temperatura, aumentando significativamente o comprimento da parte aérea, bem como o comprimento, a largura das folhas e o número de nós.
O mecanismo de ação das cepas bacterianas rizosféricas promotoras de crescimento das plantas inclui principalmente o aumento da disponibilidade de nutrientes, a produção de fitormônios e a indução de resistência sistêmica. Isolados bacterianos da rizosfera mostraram-se mais eficientes na produção de AIA do que os isolados do solo, estando associados ao aumento do crescimento das plantas (Antil et al., 2021).
4. Projeções e expectativas futuras
Os desafios futuros no manejo de culturas indicam a necessidade de classificar microrganismos de acordo com suas funções específicas e tipos de cultivo, a fim de potencializar o rendimento das culturas e otimizar a utilização dos nutrientes do solo. Além disso, torna-se imprescindível a avaliação de novos microrganismos, cepas e nichos ecológicos, com o objetivo de identificar funções específicas e aprimorar o processo agrícola.
Um aspecto de grande relevância é a busca por cepas mais resistentes, capazes de sobreviver em condições ambientais adversas, ou seja, a seleção de microrganismos com estirpes adaptadas a ambientes estressantes. Dessa forma, é fundamental incentivar o desenvolvimento de pesquisas que avaliem a eficiência dos microrganismos, suas especificidades, bem como a descoberta de novas cepas para aplicação agrícola, incluindo o estudo das melhores práticas de manejo para esses organismos.
1Universidade Federal de Santa Maria (UFSM), Santa Maria, Rio Grande do Sul, Brasil.
2Autor para correspondência: martin.ufsm@gmail.com
Referências
ANDREOTE, F. D.; CARDOSO, E. J. B. N. Introdução à biologia do solo. In: CARDOSO, E. J. B. N.; ANDREOTE, F. D. Microbiologia do Solo. Piracicaba: Editora da Universidade de São Paulo, 2016, p. 9-22.
ANTIL, S. et al. On the potential of Bacillus aryabhattai KMT-4 against Meloidogyne javanica. Egyptian Journal of Biological Pest Control, v. 31, n. 1, p. 1-9, 1, 2021
ARSHAD, M,: Frankenberger W. T, 1991. Microbial production of plant hormones. Plant and Soil, v. 33, p. 1-8, 1991.
BARBOZA, L. G. A. et al. Para além do discurso ambientalista: percepções, práticas e perspectivas da agricultura agroecológica. Ambiência. v. 8. n. 2. p. 389-401, 2012.
BORKERT, C. M. Nutrientes minerais na biomassa da parte aérea em culturas de cobertura de solo. Pesquisa Agropecuária Brasileira, v. 38, p. 143-153, 2003.
BRAOS, L. B. et al. Organic phosphorus fractions in soil fertilized with cattle manure. Revista Brasileira de Ciência do Solo, v. 39, n. 1, 2015.
CARVALHO, A. M. et al. Forms of phosphorus in an oxisol under different soil tillage systems and cover plants in rotation with maize. Revista Brasileira de Ciência do Solo, v. 38, n. 3, 2014.
CHULZE, S. N. Agentes de biocontrole baseados em microrganismos para reduzir o impacto de fungos patogênicos e toxicogênicos. Revista Argentina de Microbiologia. v. 55, n. 1, 2023.
CONTE, E.; ANGHINONI, I.; RHEINHEIMER, D. S. Fósforo da biomassa microbiana e atividade de fosfatase ácida após aplicação de fosfato em solo no sistema plantio direto. Revista Brasileira de Ciência do Solo, v. 26, p. 925-930, 2002.
COUTINHO, F. P.; FELIX, W. P.; YANO-MELO, A. M. Solubilization of phosphates in vitro by Aspergillus spp. and Penicillium spp. Ecological Engineering, v. 42, p. 85-89, 2012.
FUENTES‐RAMÍREZ, A. et al. Short-term response of soil microorganisms, nutrients and plant recovery in fire-affected Araucaria araucana forests. Applied Soil Ecology, v. 131, p. 99-106, 2018.
GATIBONI, L. C. et al. Fósforo da biomassa microbiana e atividade de fosfatases ácidas durante a diminuição do fósforo disponível no solo. Pesquisa Agropecuária Brasileira, v. 43, n. 8, p. 1085-1091, 2008.
GATIBONI, L. C. et al. Phosphorus forms and availability assessed by 31P-NMR in successively cropped soil. Communications in Soil Science and Plant Analysis, v. 36, p. 2625-2640, 2005.
HAWKES, C. V. et al. Fungal community responses to precipitation. Global Change Biology, v. 17, n. 4, p. 1637–1645, 2010.
JOSEPH, B. et al. Characterization of plant growth promoting rhizobacteria associated with chickpea (Cicer arietinum L.). International Journal of Plant Production, v. 1, n. 2, p. 141-152, 2007.
KHALID, A.; ARSHAD, M.; ZAHIR, Z. A. Screening plant growth‐promoting rhizobacteria for improving growth and yield of wheat. Journal of Applied Microbiology, v. 96, n. 3, p. 473-480, 2004.
KLOEPPER, J. W. et al. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature, v. 286, n. 5776, p. 885-886, 1980.
KLOEPPER, J. W.; LIFSHITZ, R.; ZABLOTOWICZ, R. M. Free-living bacterial inocula for enhancing crop productivity. Trends in Biotechnology, v. 7, n. 2, p.39-44, 1989.
KONIETZNY, U.; GREINER, R. Molecular and catalytic properties of phytate-degrading enzymes (phytases). International Journal of Food Science and Technology, v. 37, p. 791-812, 2002.
LAJTHA, K.; HARRISON, A. F. Strategies of phosphorus acquisition and conservation by plants species and communities. In: TIESSEN, H. (Ed.). Phosphorus in the global environment: transfers, cycles and management. Chichester: J. Wiley, 1995.
LEITE, J. N. F. et al. Frações orgânicas e inorgânicas do fósforo no solo influenciadas por plantas de cobertura e adubação nitrogenada. Pesquisa Agropecuária Brasileira, v. 51, n. 11, p. 1880-1889, 2016.
MAHDI, S. S. et al. Phosphorus availability issue- its fixation and role of phosphate solubilizing bacteria in phosphate solubilization. Research Journal of Agricultural Sciences, v. 2, n.1, p. 174–179, 2011.
MARIN, F. R. et al. Intensificação sustentável da agricultura brasileira: Cenários para 2050. Revista de Política Agrícola. n. 3. 2016.
MARTINAZZO, R. et al. Fósforo microbiano do solo sob sistema plantio direto afetado pela adição de fosfato solúvel. Revista Brasileira de Ciência do Solo, v. 31, n. 3, p. 563-568, 2007.
MAURYA, B. R. et al. Potassium-solubilizing microorganism in evergreen agriculture. In: MEENA V. S. et al. (eds) Potassium Solubilizing Microorganisms for Sustainable Agriculture. Springer, 2016.
NUNES, D. A. D. Mineralização de fósforo orgânico em solos sob leguminosas florestais, floresta secundária e pastagem. Dissertação – (Mestre produção vegetal) - Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, Rio de Janeiro, 2011.
OLANIYAN, F. T. et al. The use of soil microbial potassium solubilizers in potassium nutrient availability in soil and its dynamics. Annals of Microbiology, v. 72, n. 1, 2022.
OLIVEIRA, L. B. et al. Formas de fósforo no solo sob pastagens naturais submetidas à adição de fosfatos. Revista Brasileira de Ciência do Solo, v. 38, p. 867-878, 2014.
PRANDO, A. M. et al. Coinoculação da soja com Bradyrhizobium e Azospirillum na safra 2021/2022 no Paraná. Circular técnica. 2022.
RAWAT, P. et al. Phosphate-solubilizing microorganisms: mechanism and their role in phosphate solubilization and uptake. Journal of Soil Science and Plant Nutrition, v. 21, n.1, p. 49-68, 2021.
RYU, R. J.; PATTEN, C. L. Aromatic amino acid-dependent expression of indole-3-pyruvate decarboxylase is regulated by TyrR in Enterobacter cloacae UW5. Journal of Bacteriology, v. 190, n. 21, p. 7200-7208, 2008.
SANTOS, D. R.; GATIBONI, L. C.; KAMINSKI, J. Fatores que afetam a disponibilidade do fósforo e o manejo da adubação fosfatada em solos sob sistema plantio direto. Ciência Rural, v. 38, n. 2, p. 576-586, 2008.
TAIZ, L.; ZEIGER, E. Fisiologia e Desenvolvimento Vegetal, 6th Edn. Porto Alegre: Artmed, 2017.
WANG, C. et al. Phosphorus solubilizing microorganisms: potential promoters of agricultural and environmental engineering. Frontiers in Bioengineering and Biotechnology. v. 11, p. 1181078, 2023.
ZAMUNER, E. C.; PICONE, L. I.; ECHEVERRIA, H. E. Organic and inorganic phosphorus in Mollisol soil under different tillage practices. Soil and Tillage Research, v. 99, p. 131-138, 2008.